図1.ツノヤハズ類のルール
1996年12月
物性研究,68(5):706-709
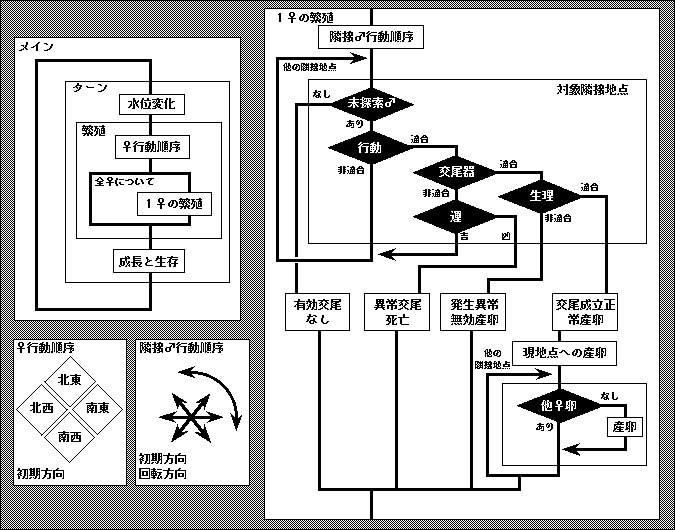
この人工生命に特徴的な事の第一は,交雑に関与する3形質の状態により,各段階の隔離機構が働く事である.生殖的隔離機構の段階の区分については各種 [2] の意見があり,細かく区分して 12 の段階が区別できる.こうちの主要な部分をを3つの隔離機構(行動的,形態的,配偶子的)に集約し,システムに組み込んだ.すなわち,1) 配偶行動形質が大きく相違すれば(つまりある許容差異よりも大きな差があれば)配偶相手として認識されず,2) 交尾器形態が大きく相違すれば正常な交尾が行われず死亡する危険が生じる.これらをクリアした上で 3) 生理的形質の差により配偶子的隔離が機能した場合,発生できない無効な卵を産むことになる.
この人工生命の特徴としてもう一点,(環境への)適応を選択の要因から排除している点がある.個体の形質状態は生存率に影響しない.個体が状況に有利/不利となる要因としては,隣接する他個体との交配に関する点のみ(個体群内での適応)である.
産下された正常卵が成虫まで生存する可能性は,地点ごとにあらかじめ決めた生存率に従うものとする.生存率の平面的な分布をマップとして与えることにより各種の分布障壁を設定,解除する.各地点が6つの隣接地点をもつ(ヘックス形式).
マップを作製するための予備的な実験として,生存および移住に必要な生存率を調査した.十分な面積をもつ場合にはヘックス単位の生存率が 0.6 程度あれば長期にわたっての生存が可能であるが,半径5ヘックス程度の狭い面積の場合には 0.75 程度が必要になる.また 0.55 程度の生存率の場所であれば,そこを経由した分布の拡大が可能である.
種は相互に交配する自然個体群のグループで,他のこのようなグループと生殖に関して隔離されたもの.
「個体群」に基礎を置くこの表現は「個体」に基礎を置く次の表現に書き換える事ができる.
2個体が直接または間接に共通子孫個体をもちうるとき,その2個体は同種である.そして種は互いに同種である個体からなる最大の集合である.
この定義はツノヤハズ類にも適用できるばかりではなく,1) 交雑の可能性について悉無的に決定でき,2) 全個体について網羅的に調査できるため,この人工生命においては現実の生物よりもむしろ厳格な適用が可能となる.
実験1.異所的種分化の再現.分布域に地理的障壁が生じることによって隔離された2群が時間を経て別の種になるシナリオは,異所的種分化 (allopatric speciation) とよばれる.この状況を地峡の断続によって再現した.3形質の許容差異各5に対して突然変異率 0.01 の状況で,二ヶ所でくびれた島に生息する約 280 個体の個体群に対して,10,000 世代から 20,000 世代まで地峡部を切断したところ,切断後 1,000 世代までに,すでに2種に分化した,この2種は交尾器形態によって機械的に隔離されているのみならず,外見的にも色彩によって十分に識別され得るものである.
実験2.地理的状況の変化を仮定しない異所的種分化.典型的な異所的シナリオにおいては明確な隔離障壁の成立と消失を想定しているが,緩やかな障壁が存在し続ける場合にも同等の効果がある.すなわち,2つの好適な生息地のあいだに比較的生存率の低い地域が存在する場合である.片方の適地が空白である時や,同種が生息している時には,移住が行われるが,別種が生息している時には互いに移住が困難となる.

好適な生息地である2つの盆地を低い峠が隔てており,片方の盆地にのみ個体群が存在する状況を設定し 100,000 世代まで観察した.個体群は 2,100 世代以内に峠を越えて分布を拡げ,その後 5,100 世代には2種に分化した.この時点で両種は配偶行動の違いによる生殖的隔離が成立しており,外見的にも識別可能であった.
その後,14,800,40,800,42,100,73,800,94,600 世代に峠を越えての分布拡大が見られ,そのうち2例は,片方の種が絶滅した空白地への進入であった.他の3例のうち一例では進入個体群が絶滅,二例では在来種が絶滅した.その場合にも絶滅後 800 世代以内に種分化が成立し,全ての場合で絶滅後 7,000 世代以内には2種併存の状況に戻った.
弱い障壁を隔てて機会的な絶滅や進入が起こり得るこの状況は,安定な2種併存の状態を繰返し作り出す,その意味で一種の復元能力がある.すなわち,一定の地理的条件が,近似種の分布状態に一定のパターンを繰返し作りだしうる.
一般に,異なる生物群で,それぞれの近似種間に共通した異所的分布パターンが見られた場合,共通の成立過程(地誌的変化)を想定する事が多い.しかし,この例に示されるように,弱い分布障壁を含む一定の地理的基盤の上で,共通の分布パターンが時を違えて繰返し生じうる.従って,共通の分布パターンは必ずしも共通の成立過程を示唆するものではないと考えられる.
実験3.側所的種分化.2つの山を鞍部がつなぐ唖鈴形の地形において,山と鞍部の生存率の組み合せを3通りの条件(1, 2, 3 高度 0, 1, 2, 3 の生存率が,それぞれ 1: (1, 1, 0.72, 0.68); 2: (0.68, 0.68, 0.68, 0.68); 3: (0.68, 0.68, 0.66, 0.52))で,種分化の成否を調べた.
その結果,高度によって生存率に格差がある2つの条件 1 と 3 で2種の側所的共存が定常的に見られた.片方が絶滅した場合にも繰返し種分化が起こり,2種共存の状態に戻る.
このように,連続した分布を保ちながら,比較的生存率の低い地域を境界として種分化がおこる現象は,典型的な側所的種分化 (parapatric speciation) である.
また,地域による生存率の格差がない条件 2 では全土同一種であった.このことから,生存率そのものではなく,生存率の格差が側所的種分化の要因となることが示された.
実験4.異所的種分化未遂による交雑帯.実験3の条件 1 と 3 において繰返し見られる側所的種分化の過程では,外見的に差が発達した2群のあいだに混血個体が生じる,いわゆる交雑帯 (hybrid zone) がしばしば観察された.
現実の生物において交雑帯の存在は,異所的種分化が未遂におわった二群が二次的に接触して生じていると説明される事が多い [5].そこで,実験1の途中の段階で地理的隔離障壁を取り除き,異所的種分化を不成立に終らせることによって,交雑帯が生じる状況を再現した.
適切な時期に海峡を取り除くことで,混血個体がみられ,交雑帯を生じたが,その存在期間は短く,100 世代以内に両群の差が不明瞭になるとともに,交雑帯も消失した.
異所的種分化の未遂によって交雑帯が成立する条件として障壁が消失するタイミングが適切であることが必要であるうえ,その存続期間も短いことから,一般には,交雑帯は側所的種分化の途中段階と解釈するのが妥当であると考えられる.